

Thierry HOQUET : Nous allons reprendre la deuxième partie de la journée. Ce matin, nous avons écouté trois très belles interventions. Nous n’avons pas rendu hommage à Olivier BROSSEAU, très grand spécialiste des créationnismes. Il n’a pas eu l’occasion de prendre la parole. Merci d’être avec nous aujourd’hui.
Nous avons un programme de 3 interventions pour cet après-midi.
Quoi de neuf dans le domaine ?
Avec Sarah SAMADI, Professeure du MNHN
Bonjour. On m’a donné comme sujet : « Quoi de neuf dans le domaine ? », j’ai donc réfléchi à ce que je pourrais illustrer de nouveau et d’intéressant dans la théorie de l’évolution. Je me suis basée sur une réflexion, que nous avons menée depuis assez longtemps avec un collègue de Montpellier, Patrice David. J’aurais pu choisir beaucoup d’autres thèmes, mais j’ai choisi d’aborder la question des relations entre l’évolution et la biologie du développement.
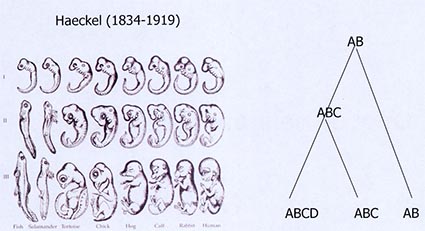
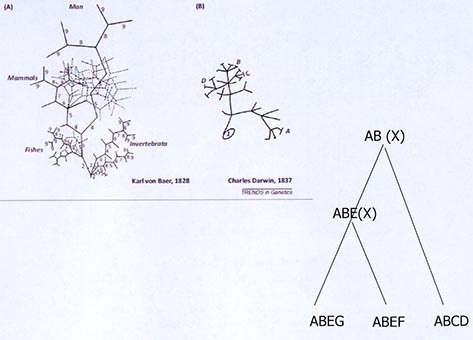
Cette diapositive illustre le fait que le lien entre l’évolution et le développement est déjà très présent dans l’œuvre de Darwin. Ici, vous voyez une figure issue des travaux de Haeckel (1834-1919), un contemporain de Darwin, qui a largement contribué à la vulgarisation des idées de Darwin. Il s’intéresse à l’embryologie comparée. Cette planche illustre les comparaisons des différents stades de développement d’un individu de différentes espèces. Il établit une comparaison des différents stades de larves au sein d’un organisme, mais aussi la comparaison des larves de différentes espèces.
Des théories, connues sous le nom de théories de la récapitulation, présentent l’idée qu’au cours de l’évolution, on complexifie le développement en ajoutant des nouveaux éléments dans ce programme qui va de l’œuf à l’adulte. À droite, cette idée est représentée sous la forme d’un arbre, où la séquence ancestrale est désignée par A et B, deux stades. Dans une des deux lignées apparaît un nouveau stade (C), partagé par les deux dernières branches. Ce schéma vous présente en quelque sorte un résumé des théories de la récapitulation. Cela fait partie des éléments pris en compte dans l’approche comparative de Darwin pour étayer les ressemblances entre les organismes, la ressemblance avec modification, etc.
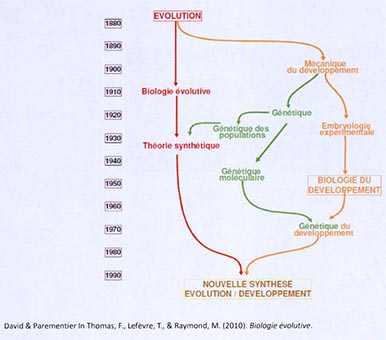
Cette figure est tirée d’un chapitre écrit par Patrice David et Marie-Laure Parmentier dans l’ouvrage collectif, Biologie évolutive. Cette figure résume l’histoire de la biologie du développement et de différents champs disciplinaires depuis la publication de la théorie darwinienne, avec une grande branche (rouge), qui est le développement de la biologie évolutive, en lien direct avec la théorie darwinienne, et en parallèle 2 autres domaines qui se développent plus ou moins indépendamment : la génétique et la biologie du développement. Au XIXe siècle, la biologie du développement, indépendamment de la théorie darwinienne, va s’intéresser à l’embryologie expérimentale, puis à la génétique du développement, un champ que l’on appelle aujourd’hui l’« évo-Dévo ». Ces travaux s’intéressent à reconsidérer ensembles les questions évolutives et celles de la biologie du développement.
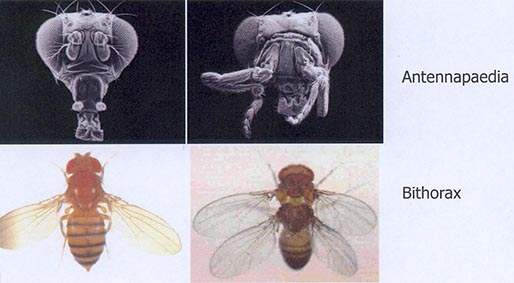
Cette nouvelle approche, qui arrive à la fin du XXe siècle est illustrée ici par les mutants de développement de la drosophile. Vous voyez ici le mutant appelé antennapaedia, c’est-à-dire une mutation génétique, qui aboutit à fabriquer des pattes à la place des antennes ; puis le mutant bithorax. Une drosophile a normalement une tête, un thorax et un abdomen. Ce mutant a deux thorax. Ces éléments arrivent dans les années 90 pour alimenter ces nouveaux débats. Ces mutants ne sont pas viables. On crée du polymorphisme pour un caractère, ce qui nous permet de comprendre quelque chose sur le développement ; ce polymorphisme, n’existe pas à l’état naturel dans les populations, car ils ne sont pas viables. Si ces mutants renseignent sur la structure génétique qui explique le développement ils ne permettent pas directement de comprendre la mise en place au cours de l’évolution de la séquence complexe qui aboutit à un organisme adulte.
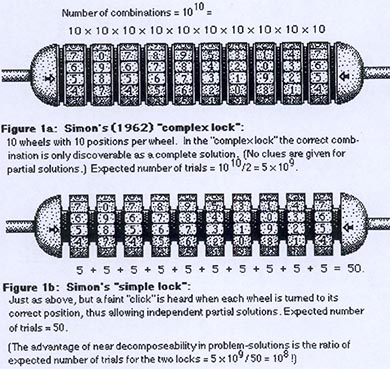
Cette grande question de l’évolution-développement est illustrée ici par ce petit système de verrous. Nous disposons d’un système fonctionnel de caractères et de stades. Il n’y a qu’une seule solution possible. Comment arrive-t-on, au cours de l’évolution, à la mise en place de cette solution, si nous n’avons pas le droit à des essais et à des erreurs (c’est-à-dire si les solutions partielles comme les mutants de la drosophile ci-dessus ne sont pas viables) ? La question est donc : Comment mettre en place une séquence complexe au cours de l’évolution ?
Ces études sur les mutations de développement, que j’ai illustrées par les mutations chez la drosophile, ont permis de décortiquer la mécanique génétique du développement. Cela a permis d’illustrer quels gènes s’exprimaient, à quel endroit, dans la larve et aboutissaient à construire ce « plan » de la drosophile.
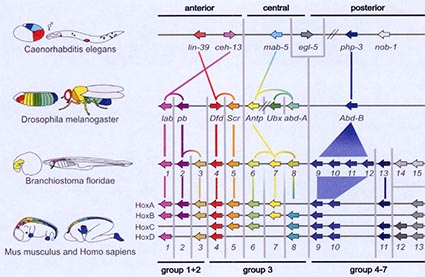
Des études comparatives sur d’autres organismes, dont les mammifères, ont mis en évidence toute une série de gènes, qui sont communs à tous ces organismes. Ces gènes régulent la façon de s’exprimer de d’autres gènes et expliquent les séquences de développements de chacun de ces organismes. De plus, il est possible de comparer ces gènes entre les différents organismes étudiés et ainsi de retracer leur évolution par une série de duplications ou de pertes, illustré dans le schéma ci-dessus. Cette reconstruction permet de proposer des modèles d’évolution qui expliquent la diversité des « plans » de ces organismes.
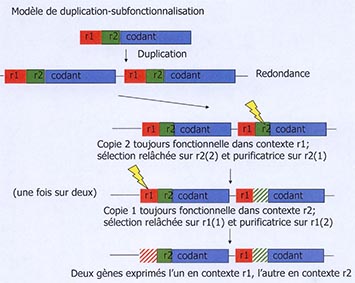
C’est le modèle de la duplication-sub-fonctionnalisation. Il est extrêmement important pour comprendre le processus de mise en place de ces gènes. Le schéma illustre ce processus. Un gène codant (en bleu) présente 2 régulateurs : le régulateur R1, qui permet son expression à un certain stade de développement de l’organisme et un autre régulateur, R2, qui permet son expression à un 2e stade. Par hasard, une duplication arrive. Le fragment d’ADN, qui code pour cette unité opérationnelle, est doublé. Cela signifie que nous avons 2 fois les outils pour permettre un certain type de fonction, ce qui permet un relâchement de la sélection naturelle. Ainsi, si une mutation arrive dans une des 2 copies, ce n’est pas grave puisqu’elle dispose d’une réplique.
Or, les mutations peuvent aussi se produire dans les parties régulatrices. Si par exemple, une mutation arrive dans la zone verte (R2 de la 2e copie), le gène n’est plus exprimé que dans le contexte R1. J’ai maintenant une 2e situation avec un gène toujours fonctionnel en R1 et R2, mais un 2e seulement fonctionnel dans R1. Cela relâche la pression de sélection cette fois sur la partie R1. Si ces 2 événements arrivent, 2 copies d’un même gène, qui avaient donc une même fonction, vont être spécifiques dans 2 contextes différents. Ces 2 fonctions vont pouvoir ensuite évoluer indépendamment et donc s’affiner et changer.
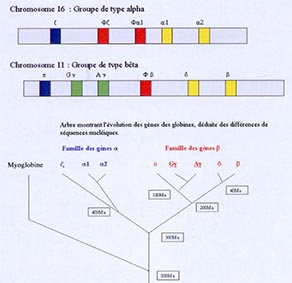
Ce modéle est ici illustré par le cas d’une famille dite multigénique : celle des globines. Les gènes, qui codent pour les globines, peuvent se diviser en plusieurs sous-familles, correspondant à différentes duplications, se déroulant au cours de l’histoire de cette lignée, avec une diversification des fonctions, qui permet que certaines formes soient exprimées chez l’embryon et d’autres chez l’adulte. Ainsi, dans le placenta, la globine fœtale a une meilleure affinité pour l’oxygène que la globine maternelle, permettant le transfert de l’oxygène depuis la mère vers l’enfant. Les globines sont des protéines à l’origine du transport de l’oxygène dans le sang. Ce système de duplication amène l’existence de plusieurs molécules différentes, permettant d’affiner les fonctions. Ce type d’explications permet de comprendre la façon, dont les développements complexes ont pu se construire au cours de l’évolution : par ces systèmes de duplication, divergences, éventuellement pertes de certaines copies.
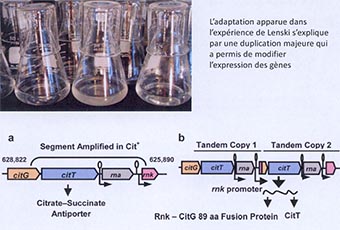
Vous n’étiez pas tous présents la semaine dernière, mais j’avais abordé une expérimentation, visant à comprendre de façon expérimentale l’évolution. Dans cette expérimentation, on utilisait des populations de bactéries dans un milieu contrôlé avec un certain nombre d’éléments nutritifs utilisables par la bactérie ou simplement présents et non utilisés. Au cours de cette expérience, on montrait que la sélection naturelle agissait sur les mutations, qui arrivaient par hasard, avec une augmentation du succès reproducteur de ces bactéries. Tout à coup, dans une lignée de cette expérience, apparaît un élément étonnant : une nouvelle lignée de bactérie exploitant un élément du milieu, le citrate, présent depuis le début, mais qu’aucune bactérie n’était capable d’utiliser. Cette expérience a pu être réalisée plusieurs fois, pour démontrer que cela n’arrivait qu’à la suite d’une série de mutations, qui devaient survenir dans un certain ordre. Parmi ces mutations, nous avons une illustration de ce phénomène de la duplication.
Un élément génétique, présent dans les bactéries depuis le début, va être dupliqué. La duplication et la recombinaison de ces séquences vont permettre de changer la façon dont cette séquence codante s’exprime ou non, ajoutée à d’autres mutations pour expliquer l’apparition de cette nouvelle adaptation. Nous sommes donc dans un système beaucoup plus simple (une bactérie), mais au final, une fonction complexe apparaît.

Ce même groupe de chercheurs, qui travaille sur les bactéries, s’est aussi intéressé à l’émergence de ce type de traits complexes, de façon informatique. Ces recherches permettent, également grâce à des processus de mutation et de duplication, d’aboutir à des fonctions complexes. Il s’agit d’un autre type d’approche expérimentale, qui appuie l’intérêt de ce type de raisonnement et de ces mécanismes, par une approche analogique.
Ce schéma montre que les modalités de développement n’ont pas lieu uniquement par ajouts. C’est-à-dire que le développement ne se résume pas à une séquence simple allant vers une séquence de plus en plus complexe avec l’ajout d’éléments, mais il y a également une variation possible de ces différents éléments. Si je regarde la dernière ligne (une variation des organismes actuels de l’arbre phylogénétique), tous les organismes présentent 4 stades. Les divergences apparaissent au fur et à mesure dans les différents stades. Il n’y a pas moins de stades chez les lignées ayant divergé depuis plus longtemps. Les données récentes de la recherche alimentent et permettent de remettre à jour l’intérêt de ce type de raisonnement, datant du XIXe siècle.
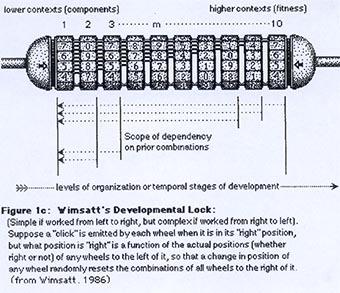
Pour conclure, on n’aboutit pas à une combinaison d’un seul coup. Un système progressif amène des séquences, qui se contraignent les unes les autres. Toute la difficulté que tout cela illustre est qu’en biologie du développement, de façon expérimentale, on crée des polymorphismes, qui n’existent pas dans les populations naturelles. On met donc en évidence le rôle de tel ou tel gène dans ce développement, mais ce n’est pas une réelle indication sur la mise en place de ce mécanisme au cours de l’évolution et notamment de la manière dont la sélection naturelle a pu agir pour trier les variants au sein des populations.
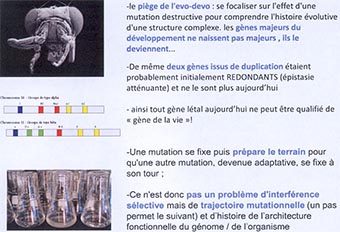
Un des pièges de ce domaine de l’évolution-développement serait de se focaliser sur les effets d’une mutation destructive pour comprendre l’histoire évolutive. Ces gènes majeurs du développement ne naissent pas majeurs. Ils le deviennent par la suite. Ainsi, les 2 gènes issus d’une duplication sont probablement redondants au départ, mais par le jeu des mutations et du relâchement de la sélection naturelle liée à cette redondance on peut comprendre pourquoi ils ne le sont plus aujourd’hui. À l’origine, une redondance permet l’apparition d’un polymorphisme et l’action de la sélection naturelle.
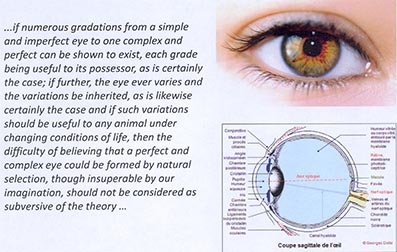
Il y a apparition d’une mutation, pouvant être une duplication, qui se fixe et prépare le terrain pour une autre mutation, qui elle-même devient adaptative. Il y a donc une trajectoire historique des mutations et de l’action de la sélection naturelle qui explique l’architecture fonctionnelle actuelle du développement d’un organisme ou d’un caractère donné. J’ai sélectionné une citation de Darwin à propos de la formation de ces mutations complexes qu’il illustré par le cas de l’œil. Il avance un mécanisme expliquant l’apparition de telles mutations : des variations intéressantes à un moment donné pour des fonctions qui ne sont pas forcément celles associées à la fonction finale de cet organe.
Les 20 dernières années ont connu de nombreux questionnements sur la base de ces recherches sur le développement et les variations : faut-il repenser la théorie de l’évolution ? Faut-il partir d’un autre type de schéma que variations, sélection naturelle et tri aléatoire ? Il est également important de définir si, toutes les variations héritables sont déterminées par des molécules d’ADN. Il s’agit d’un des éléments importants du débat actuel. Est-ce que seules les variations des organismes, qui s’expliquent par une variation de l’ADN, sont importantes pour comprendre l’évolution ? Certains répondent que finalement la sélection autour des caractères liés à l’ADN n’est pas l’essentiel. Les variations de d’autres natures et la construction des organismes à travers le développement seraient selon eux plus importantes pour expliquer l’évolution.

L’ADN est une molécule, transmise lors de la reproduction quasiment telle quelle, avec des caractères, qui ne vont pas varier au cours de la vie de l’organisme. Nos parents nous transmettent un ADN que nous retransmettons quasiment tel quel à nos descendants, avec quelques petites mutations. Or d’autres traits des organismes peuvent être héritables, mais ne sont pas déterminés par l’ADN. C’est le cas de compétences telles que la capacité à faire du feu. Je peux transmettre cette compétence à mes enfants, qui la transmettront à leur tour à leurs enfants. Ce trait n’est pas expliqué directement par la génétique, mais transmis de façon héritable par apprentissage.
Par exemple, certains oiseaux vont être capables d’exploiter différentes ressources en fonction de leur environnement. Cette plasticité n’est pas forcément déterminée génétiquement. Ce qui est déterminé génétiquement, c’est la capacité à explorer différentes ressources présentes dans l’environnement. Pourtant, dans un environnement donné doté d’une ressource particulière, certains oiseaux vont apprendre à l’exploiter et transmettre à leur descendant les connaissances pour exploiter également cette ressource. Si par hasard, un descendant a une « mutation », lui permettant d’exploiter cette ressource, même s’il ne l’a pas acquis de ses parents, cette mutation a de grandes chances de se fixer sur la population, qui vit sur cette ressource et donc à fixer le destin de cette variation, de façon définitive dans cette lignée.

Ce type de raisonnement est actuellement beaucoup développé et s’intéresse aux variations en général, héréditaires, pas forcément liées à l’ADN. Du temps de Darwin, on ne disposait pas de très bonne théorie de la variation, ni du développement, mais Darwin a tout de même développé une théorie de l’évolution. Les recherches au cours du XXe siècle, jusqu’à aujourd’hui, ont permis de mieux comprendre les bases génétiques et moléculaires de l’hérédité, et de mieux comprendre le développement des organismes. Nous nous rendons compte de nos jours que l’absence de théorie précise de l’hérédité chez Darwin n’est finalement pas problématique. L’essentiel est bien de disposer d’un système variable, où la variation peut être transmise à ses descendants via un processus que nous appelons l’hérédité.
Je voulais terminer là-dessus.
Merci de votre attention.
Applaudissements
Thierry HOQUET : Merci Sarah SAMADI, pour ces clarifications. Y a-t-il des questions ?
Dominique GUILLO : J’aimerais préciser quelque chose sur l’hérédité, qui n’est pas génétique. Il me semble important de souligner que l’hérédité culturelle est considérée par tous les théoriciens qui la défendent, comme une hérédité lamarckienne. Elle est en effet orientée par des choix.
Sarah SAMADI : Pas forcément. Elle peut être aléatoire. Je pense par ailleurs que ce débat peut être assez vaste. Je crois que l’important est que Darwin n’a pas défini de réelle théorie de la transmission héréditaire. Ces études et ces questionnements démontrent bien qu’il peut exister différentes formes de variation et d’hérédité, mais que, dans tous les cas, le processus de sélection naturelle et de dérive est une explication puissante pour comprendre l’apparition d’adaptations au cours de l’évolution, même si le pas initial de la variation peut être du registre de l’hérédité non génétique. La grande différence d’une hérédité génétique et d’une hérédité liée à l’apprentissage culturel est que la pérennité dans le temps n’est pas la même que celle qui est liée à l’ADN.
Dominique GUILLO : Il me semble qu’une autre différence est invoquée dans cette littérature. Il n’y a pas d’indépendance entre les causes de la variation et les causes de la sélection. C’est-à-dire que la raison pour laquelle un trait se diffuse est la raison même pour laquelle il apparaît. L’évolution est guidée.
Thierry HOQUET : J’aimerais formuler à ce propos une objection classique tirée des traditions évoquées ce matin. En effet, nous avons parlé ce matin des théories des lamarckiens français, en particulier d’Albert Vandel. Ce dernier considère que l’arbre de l’évolution comporte des paliers. Or, pour lui, ces paliers ne constituent pas l’essentiel de la théorie de l’évolution, mais que l’essentiel est plutôt les changements de paliers.
Or, j’ai l’impression que les exemples que tu as choisis sur l’évolution moléculaire se situent toujours dans cette zone. L’expérience des bouteilles de Lenski en est un bon exemple. L’apparition d’une nouvelle capacité n’est pas un changement d’organisation aussi important que ce changement de palier. C’est une objection qui a couru dans la biologie française au XXe siècle. Les mutations ne concernent pas ce grand changement de plan d’organisation. Comment répondez-vous à cela aujourd’hui ?
Sarah SAMADI : Personnellement, je considère que le changement, qui se produit dans l’expérience de Lenski, est un grand changement. En effet, cette mutation paraissait impossible au moment où l’expérience a été constituée. Cela veut bien dire que, puisque cet élément était hautement improbable, il est qualitativement vraiment très différent. Il est toujours quelque peu subjectif de définir à quel moment il y a variation ou non. Dans un contexte contrôlé et une constitution génétique bien connue, apparaît quelque chose qui, du point de vue microbiologique paraissait complètement improbable.
Laurent LOISON : Je me fais l’avocat du diable en suivant la question de Thierry HOQUET. Un des gros problèmes de la compréhension de la théorie de l’évolution est l’articulation entre la microévolution et la macroévolution. Il me semble que la réponse, que vous formulez à cette question, est quelque peu hors sujet, puisqu’il s’agit vraiment d’une question théorique. Comment articule-t-on le temps court au temps long à l’intérieur de la théorie de l’évolution ? Cette question est très difficile et a été résolue d’une certaine manière au moment de la théorie synthétique de l’évolution par ce qu’on appelle le principe d’extrapolation.
Toutefois, elle est remise sur le devant de la scène par la théorie de Stephen Jay Gould, des équilibres ponctués. C’est notre manière de faire le lien entre micro et macroévolution. Du point de vue des enjeux éducatifs, il est extrêmement compliqué de confronter les gens à cette distinction, puisqu’énormément de personnes sont susceptibles d’accepter le darwinisme à l’échelle de la macroévolution. Or ils vont découpler cela assez spontanément de ce qui se produit à l’échelle de la macroévolution. C’est ce qui se passe à l’heure actuelle, quand on enseigne cela à des élèves. Historiquement, comme le disait tout à l’heure Thierry HOQUET, on retrouve cela à l’échelle de l’histoire des sciences aux XIXe et XXe siècles.
À mes yeux, c’est plus qu’une question empirique. Vous avez répondu sur le terrain empirique en soulignant que, dans cette expérience, l’intérêt était lié à la très faible probabilité de la mutation, qui en faisait un grand rameau. Cependant, pour moi, la question dépasse largement cet exemple et reste encore une question théorique largement ouverte à l’intérieur de la théorie de l’évolution, faisant appel à la structure de la théorie de l’évolution. Cela reste un point passionnant, ouvert et évidemment très compliqué.
Sarah SAMADI : Je pense que ce type d’expérimentation peut être exploité avec un raisonnement analogique pour expliquer le changement entre micro et macro-écologie. Il est important de disposer de corpus expérimentaux pour étayer la façon, dont nous pensons ce passage. Pour rejoindre ces questions, je souligne que tout le champ de l’évolution développement s’est davantage intéressé à des questions macro-évolutives à la fin du XXe siècle.
Les recherches actuelles dans un domaine en plein développement (« l’éco-évo-dévo ») s’intéressent aux polymorphismes existants dans les populations, qui peuvent être le sujet à sélection naturelle et qui, dans le fameux cadenas, permettent de comprendre comment on arrive à jouer là-dessus. Ce type d’expérimentation de Lenski, même sur des organismes très simples et sur une expérience de 60 000 générations multipliée par 12, a un rôle extrêmement important dans le pouvoir explicatif de la théorie et dans la façon, dont on va étendre ce qu’on comprend à l’échelle micro pour comprendre l’échelle macro.
Guillaume LECOINTRE : Je voudrais apporter une référence bibliographique pour ceux que l’hérédité étendue intéresserait. Un Pour la science, paru en décembre 2013, intitulé « L’hérédité sans gène », présente de nombreux exemples d’héritabilité qui n’est pas génétique, dont les conséquences ne sont par ailleurs pas nécessairement favorables au porteur. La question du lamarckisme ne se pose pas trop en l’occurrence dans ce cas.
Je voudrais apporter un complément de réponse à la question de Laurent sur la macroévolution. Malheureusement, les biologistes du développement d’aujourd’hui utilisent encore une notion à mon sens critiquable : la notion de plan d’organisation. Un plan d’organisation reste un bon outil mnémotechnique pour l’enseignement, pour retenir comment est formé un vertébré. Quand nous regardons la diversité du vivant, nous nous apercevons que les pièces du puzzle anatomique ne sont pas si rigidement reliées les unes aux autres et que le plan d’organisation est déjà, en réalité, caduc.
Si je vous parle de « plan » des vertébrés, cela présuppose que j’ai déjà à l’esprit une boîte regroupant tous les vertébrés en gommant les irrégularités. Le taxon est fixe et je fais entrer des combinaisons de caractères de force dans la boîte. Or, en phylogénie c’est l’inverse : les caractères ne sont pas reliés entre eux, par contre ils sont utilisés pour justifier la boîte qui en découle. Le plan fait passer la boîte avant sa justification, la phylogénie fait passer la justification de la boîte avant, afin de la créer. Je veux dire par là que la difficulté du passage d’un niveau micro-évolutif au niveau macro-évolutif est en partie liée au fait que j’ai rigidifié dans mon esprit un plan « annélide », puis un plan « arthropode ». Or, si vous raisonnez en phylogénéticien et non plus en plans idéalisés, comme Goethe les avait pensés, la phylogénie vous restitue la compréhension évolutive du passage entre une mosaïque de caractères et une autre : il n’y a plus de « trou » si marqué entre annélides et arthropodes, parce que la définition des groupes se traite en termes de conventions fixées sur des caractères, et non des plans. Et une bonne partie de la question de la différence entre micro et macro évolution s’estompe. En effet, la macroévolution concerne l’évolution entre les plans. Si les plans n’existent pas, la différence entre la macroévolution et la microévolution ne se pose plus, sauf dans des termes très exacts que Sarah a spécifiés.
Ainsi, il y a des moments, qui présentent des changements relativement importants dans l’organisation de l’organisme. L’importance dans les conséquences de changement n’implique pas nécessairement une macroévolution au sens où le mot a été utilisé au début du XXe siècle. J’ai, pour ma part, passé ma carrière à entendre parler de différence entre macroévolution et microévolution. Or je me suis aperçu que, bien souvent, cela résultait d’abus de langage que nous, scientifiques, commettons nous-mêmes. Tout le monde n’est peut-être pas d’accord dans la salle. En tout cas, certains événements génétiques ont des conséquences lourdes sur l’organisation de l’organisme. Quant à appeler cette organisation un plan, je suis sceptique.
Auditrice : Dans les exemples que vous avez cités, vous avez parlé d’hérédité génétique et d’hérédité culturelle. L’épigénétique est-elle un élément venu en renfort, en opposition ou en contrepoint de tout le reste ?
Sarah SAMADI : Guillaume a cité des exemples. Effectivement, il y a un certain nombre de variations tout à fait matérielles transmises d’un organisme à ses descendants, sans s’expliquer directement par l’ADN. L’épigénétique s’adresse à des aspects matériels, alors que quand nous parlons d’apprentissage, nous abordons une notion moins matérielle. L’épigénétique comprend un certain nombre de caractères variables, qui vont être transmis et qui sont souvent défavorables au porteur.
Ce type de variations touche à un champ assez actif de la recherche contemporaine, qui se penche sur la vision gène-centrée, qui aurait pu gommer d’autres aspects de la théorie de la variation. Dans le débat, certains pensent qu’il faut davantage se concentrer sur différents types de variations, en particulier la variation épigénétique. D’autres considèrent que la partie gène-centrée reste pratique et a permis la compréhension de beaucoup d’éléments. Cela n’empêche pas d’autres variations. Cela entre dans le cadre théorique sans la changer fondamentalement. Le débat, illustré par l’article que je vous ai montré précédemment, englobe ces sujets. Certains considèrent que le domaine de recherche en épigénétique bénéficie d’une approche publicitaire, mettant en valeur l’aspect fondamental de ces recherches. L’autre partie du discours est de considérer que cela entre dans la structure globale de la théorie de l’évolution, qu’il faut l’affiner, mais non révolutionner sa structure.
Thierry HOQUET : Merci Sarah.
Je cède la parole à Philippe HUNEMAN, philosophe des sciences, coéditeur du volume collectif, Les mondes darwiniens et de nombreuses publications.
Applaudissements