

L’empreinte des premières domestications sur le squelette animal,
Thomas CUCCHI (AASPE - CNRS / MNHN, Paris)
Frédéric DENHEZ : […] On passe du génotype au phénotype : « Domestication, évolution phénotypique », avec vous Thomas CUCCHI. Où êtes-vous ? Je ne vous vois pas sur mon écran. Je suis très inquiet. Non, en fait, je ne suis pas inquiet, du tout !
Thomas CUCCHI : Je suis là !
Frédéric DENHEZ : Vous êtes là, vous êtes limite en contre-jour, ce n’est pas gentil pour nous.
Voilà, c’est parfait ! Allez-y.
On a tendance à penser que la domestication se caractérise par une modification complète de la morphologie de l’animal d’origine, en fait ce n’est pas aussi évident que cela, vous allez nous le dire, on vous écoute.
Thomas CUCCHI : Je vais vous dire cela tout de suite.
Bonjour à tous,
Merci pour cette invitation.
Parler après Ludovic ORLANDO, c’est un peu comme monter sur scène après James Brown, je vais quand même essayer de vous intéresser, avec une recherche que l’on est en train de mener dans le laboratoire de biologie archéologique du Muséum, sur comment on met en évidence le début des premières domestications, à partir des vestiges archéologiques, les ossements d’animaux.
Pourquoi s’intéresser aux débuts de la domestication ? ce n’est pas moi qui le dit, mais c’est vraiment aller aux racines de l’Anthropocène, puisqu’en effet, l’une des composantes de l’Anthropocène, c’est l’érosion de la biodiversité. Or, si on observe sur ce graphique la distribution de la masse […]
Frédéric DENHEZ : On ne voit pas votre graphique, Thomas. Je propose que, si vous avez du mal avec votre écran, qu’Élodie le diffuse. Vous n’avez pas fait le partage d’écran. […]
Thomas CUCCHI : Ah, oui, je n’ai pas fait de partage d’écran. Mais qu’est-ce que je fais ! ?
Frédéric DENHEZ : Mais ça, c’est James Brown qui vous perturbe 
Thomas CUCCHI : Mais, voilà ! Attendez, j’arrive, je partage. Excusez-moi.
Frédéric DENHEZ : Vous l’êtes. Magnifique ! Bravo ! On vous écoute et on vous regarde.
Thomas CUCCHI : […] Voilà le graphique, que vous voyez maintenant, j’espère.
Frédéric DENHEZ : Très bien, merci !

Thomas CUCCHI : Parfait ! Donc, ça, c’est la distribution de la masse des mammifères terrestres, c’est issu des travaux de Vaclav SMIL, on voit qu’ils 97% de la masse des mammifères terrestres sont représentés par les humains et surtout par leurs animaux domestiques, les mammifères sauvages ne représentent plus que 3% de cette masse terrestre. Aller aux racines de la domestication, dans l’histoire évolutive de notre espèce, c’est comprendre pourquoi après trois-cent-mille ans d’évolution de l’homme comme chasseur-cueilleur une partie de l’humanité a choisi une trajectoire de la production alimentaire, qui nous a menés jusqu’aux crises écologiques qui touchent désormais toute l’humanité.
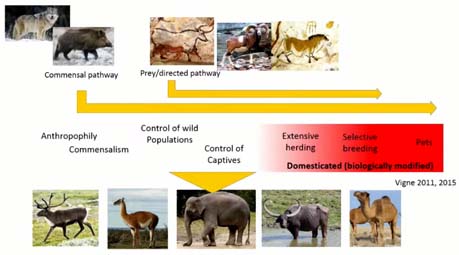
Mais aller aux racines de la domestication, c’est quand même pas une chose aisée, pour les archéologues nous sommes, tout d’abord parce qu’on travaille sur des vestiges archéologiques qui sont fragmentés, issus de contextes sens en écriture, aussi, comment vous l’a expliqué Jean-Denis VIGNE, c’est qu’allait aux origines de la domestication, c’est mettre en évidence une intensification des relations entre l’homme et l’animal, qui sont le montrés sur ce graphique, qui est un continuum sans frontières nettes ; et si l’on se contente d’une définition biologique de l’animal domestique, un animal sélectionné par l’homme, qui présente des caractéristiques génétiques, physiologiques et morphologiques différentes des populations sauvages, eh bien, on se prive de toutes ces interactions qui précèdent et qui sont le processus de domestication. Ce processus de domestication a pu durer plusieurs millénaires, être initié, abandonné, et prendre place parmi de nombreuses sociétés de chasseurs cueilleurs à travers le monde.

L’un des problèmes pour les archéologues qui étudient la domestication, est qu’ils se sont beaucoup appuyé sur ce qu’on appelle le syndrome de la domestication chez les mammifères, décrit par DARWIN à la fin du XIXe siècle. Il s’agit de cette suite de caractères morphologiques et comportementaux qui sont : le changement de la morphologie crânienne, avec le raccourcissement de la face, les oreilles tombantes, les dents plus petites, le changement de la couleur du pelage, mais aussi un comportement plus docile, la réduction de la taille du cerveau, les changements hormonaux mais aussi une augmentation des taux de reproduction. Ces critères étaient considérés comme les marqueurs absolus de l’animal domestique, depuis surtout les travaux expérimentaux du généticien Dmitry BELYAEV, qui a démontré expérimentalement, que ces caractères, ces syndromes de domestication étaient la conséquence d’une sélection de comportements dociles. Pour le démontrer, il a rassemblé une population de renards argentés, comme moi on le voit sur la photo, à peu près 30 mâles et 100 femelles, il n’a gardé pour la reproduction que les individus les moins agressifs, les plus aimables vis-à-vis de l’homme. Et, en dix générations, il a réussi à reproduire tous ces syndromes de domestique. Vous voyez là ce renard avec tous les syndromes de domestication possible, docile comme un toutou.
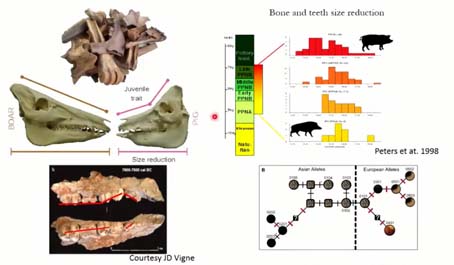
Ensuite, les archéozoologues ont utilisé un certain nombre de domestications sur le squelette, à savoir la réduction de la taille des os et des dents, le raccourcissement de la face et ses conséquences, par exemple le chevauchement des dents, lié à la malocclusion produit par le raccourcissement de la face. Et plus récemment, ils ont cherché un changement de la couleur du pelage dans le génome.
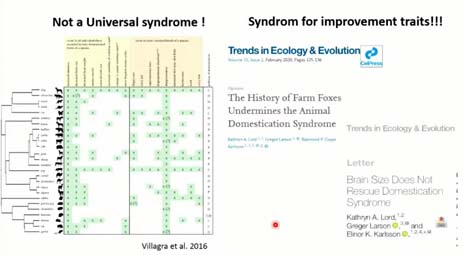
Le problème est que ces syndromes de domestication ne sont efficaces pour aller aux racines de la domestication animale, et ce pour au moins trois raisons. Tout d’abord, parce qu’il est peu vraisemblable qu’une telle sélection comportementale ait eu lieu lors des premières domestications, car l’isolement génétique qu’elle sous-tend a été démontrée comme peu probable par les travaux de génétique. Ensuite, ces syndromes, comme on voit sur ce diagramme, avec tous les animaux domestiques sur cet axe-là et tous les syndromes sur l’axe des Y, ne sont absolument pas universels, c’est ce que montrent tous ces creux blancs. Ça, c’est la deuxième chose. Plus fondamentalement, des travaux récents, qui ont scrutés les données, ont montré d’une part que l’expérimentation, sur les renards argentés, était issue d’une population qui était en captivité depuis plus d’un siècle, c’est pour cela que le résultat est extrêmement rapide. Et, de façon plus fondamentale encore, pour mettre en place ces syndromes, les comparaisons ont été faites en des populations domestiques et des populations sauvages qui n’avaient aucun lien évolutif et historique entre eux. Par exemple, ne serait-ce que pour la réduction du crâne, en général on comparaît des chevaux domestiques avec des chevaux de Przewalski, comme l’a montré Ludovic, qui n’ont pas grand-chose à voir. Ils ont comparé des moutons avec des mouflons européens. Donc, ces syndromes de domestication ne sont pas efficaces pour vraiment aller aux racines de la domestication.
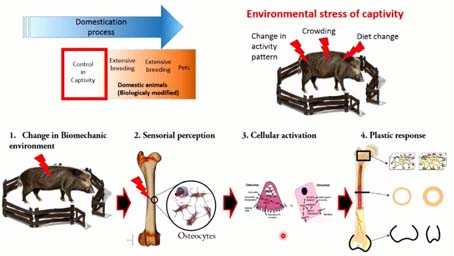
Pour essayer de lever ce verrou méthodologique, on a voulu se concentrer sur les stresses que représente l’entrée de l’animal dans l’environnement anthropogène, dans la domus, à travers le prisme de la captivité et du contrôle de la mobilité d’un animal sauvage. Cette approche nous semble un peu plus universelle permettrait d’appréhender une plus grande quantité de la première domestication à travers le temps et l’espace. Parmi les stresses environnementaux de la captivité, on s’est d’abord intéressé à la réduction de la mobilité liée à la captivité. On sait que l’anatomie osseuse, grâce à sa plasticité, peut enregistrer ce type de changements de modes de vie, parce qu’effectivement un os se modèle et se remodèle en fonction de son activité biomécanique dans son environnement, grâce à activation cellulaire qui va produire différentes structures à travers les chaînes de la structure osseuses.
Frédéric DENHEZ : Vous avez fait cela sur des sangliers, pas à la ménagerie du Muséum, pas au Jardin des plantes, l’êtes.
Thomas CUCCHI : Non, mais j’en viens.
Frédéric DENHEZ : Bon, voilà !
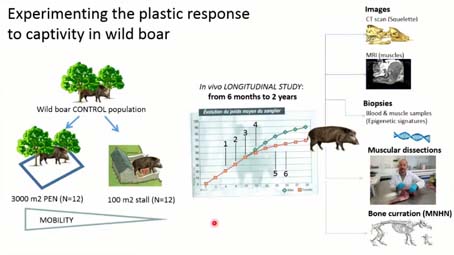
Thomas CUCCHI : J’arrive, excusez-moi, je m’étends Il fallait qu’on démontre quand même que la captivité entraînait des changements à l’’échelle de la vie de l’individu, des changements qui n’avaient avec la variation que l’on pouvait observer dans le milieu sauvage. Donc, on a fait une expérimentation sur le sanglier. Pourquoi le sanglier. Parce que le sanglier est une des rares espèces ancestrales sauvages d’une espèce domestique, qui n’est ni voie d’extinction ni éteinte, et surtout qui présente de nombreuses populations à l’échelle de l’Eurasie. Donc, on pouvait faire une expérimentation dessus. Pour contrôler un peu l’effet génétique sur la variation morphologique, on a capturé des marcassins de six mois dans une population contrôle, on l’a ensuite divisé en deux lots de façon aléatoire, au moment de la séquestration et on les a fait grandir dans deux environnements biomécanique différents, l’un dans un enclos de 3 000 m², et l’autre dans une stabulation de 200 m². et, au cours de leur croissance, jusqu’à deux ans, tous les trois mois ils ont passé un scanner et une IRM, ainsi que des biopsies, des prises de sang. Et à la fin de l’expérimentation, ils ont été disséqués, mesurés, et leur squelette fait partie maintenant des collections patrimoniales du Muséum.

Pour comparer la variation plastique liée à la captivité, à ce que l’on appelle la norme de réaction d’un sanglier dans son milieu naturel, nous avons scanné des populations de sangliers, dans différentes forêts en France, en Suisse, en Allemagne, et pour comparer la plasticité liée à la captivité face à la variation liée à la sélection des races, nous avons collecté des cochons de différents types d’élevages, plutôt extensifs, des races plus traditionnelles et des races de type industriel, plus récentes, qui ont été vraiment sélectionnées pour la croissance rapide, et la masse musculaire.
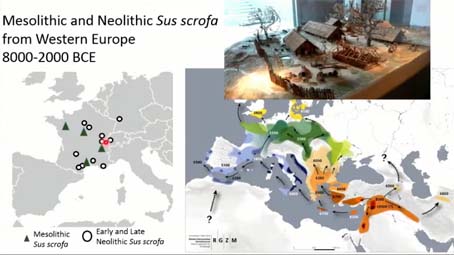
Nous avons voulu explorer ce signal de leur registre archéologique, nous avons scanné des restes archéologiques de (manque un mot, incompris), provenant de deux contextes mésolithiques, dans ces triangles-là. Le Mésolithique, ce sont les derniers chasseurs-cueilleurs d’Europe occidentale, entre 8 000 et 6 000 avant Jésus-Christ, et nous avons aussi collecté des spécimens issus de sites du Néolithique ancien jusqu’au Néolithique récent, qui sont arrivés en Europe occidentale, selon ces deux voies de diffusion, comme vous le voyez sur cette carte.
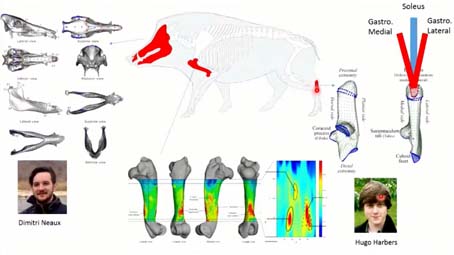
Pour l’instant, nous avons pu observer l’effet de la captivité sur le squelette sur trois éléments osseux, deux éléments des membres, qui ont été étudié dans le cadre de la thèse d’Hugo HARBERS, …
Frédéric DENHEZ : Pourquoi ces éléments osseux et pas d’autres ?
Thomas CUCCHI : Ces deux, là, parce qu’ils sont bien conservés en contexte archéologique. Pour le calcanéum, c’est aussi parce qu’il est extrêmement important dans la locomotion, il est connecté avec trois muscles extrêmement puissants, qui propulsent l’animal. L’humérus est bien conservé en contexte archéologique, surtout dans sa partie distale. Le crâne n’est pas impliqué dans la locomotion mais de par sa fonction, il va répondre aux changements de modes d’acquisition de l’alimentation, de la compacité de l’alimentation lié aux changements de mode de vie. Le calcanéum et le crâne, qui a été étudié par Dimitri NEAUX, ont été analysés en morphométrie géométrique tridimensionnelle. Pour l’humérus, on s’est plutôt intéressé à l’épaisseur de la diaphyse, avec une approche en cartographie morphométrie, où là on regarde les patrons d’épaisseur, la topographie d’épaisseur corticale, puisque cela nous donne des informations sur l’activité musculaire. Là, vous voyez les zones en rouges, ce sont les zones d’insertions musculaires.
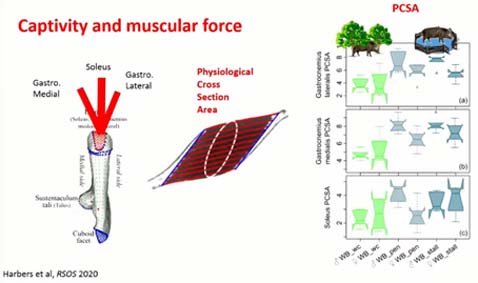
Là, on va regarder les résultats. Tout d’abord, on va regarder l’effet de la captivité sur l’activité musculaire au niveau du calcanéum. Pour cela on fait une mesure qui s’appelle la physiologecal cross section area, c’est cette section entourée en plan, qui comprend en parallèle toutes les fibres musculaires. Comme la force musculaire est liée au nombre de fibres musculaires, on a là une sorte de proxy de la force musculaire. Si l’on regarde ces trois box-plots de la variation de la force musculaire, on voit que les sangliers captifs en bleu, ont en général plus de force musculaire que les sangliers en liberté, ici en vert. C’est assez surprenant, mais on pense que la captivité a entraîné des comportements répétitifs, qui associés à une alimentation plus constante et plus nutritive a peut-être favorisé la production de muscles de type 2, plus gros et plus puissants. Cela serait la différence entre les muscles du bodybuildeur ou de sprinters et des muscles de marathoniens.
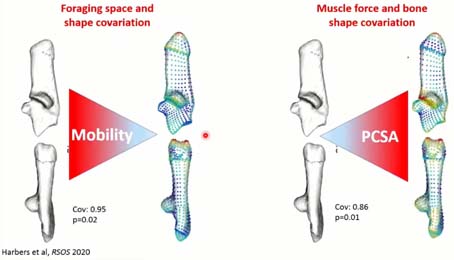
Si on observe comment la forme du calcanéum change, en fonction de la réduction de la mobilité et l’augmentation de la force musculaire, on observe que ces changements sont principalement localisés sur les zones d’insertion musculaire. La réduction de la mobilité associée à une augmentation de la force musculaire laisse une empreinte sur le calcanéum.
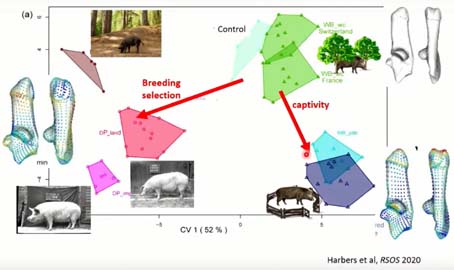
Si on compare cette réponse plastique à la sélection, sur ce graphique, on voit que la captivité entraîne une différence de variation, bien au-delà de la variation de sangliers dans leur milieu, mais selon une direction différente de la sélection qui, elle, a entraîné des phénotypes beaucoup plus jeunes robustes et épais. On pense que la sélection d’une croissance plus rapide, plus musculeuse, ou plus lourde a changé les forces musculaires, les contraintes au sol, qui ont entraîné des ajustements du calcanéum.
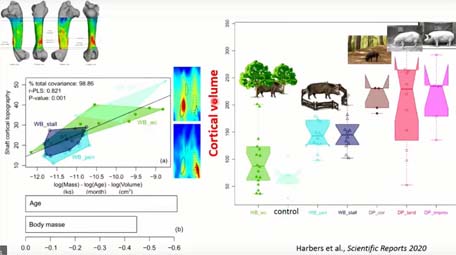
Si on passe au niveau de l’épaisseur corticale, chez les sangliers, on voit que principalement ce qui influence la topographie de l’épaisseur corticale, c’est principalement l’âge et la masse musculaire. Et si on regarde le volume cortical, on peut observer que les individus captifs semblent avoir un volume cortical plus importante que les sangliers, mais peut-être c’est parce qu’on manque d’individus plus âgés, par contre, on voit que les cochons domestiques, eux, ont un volume cortical beaucoup plus important.
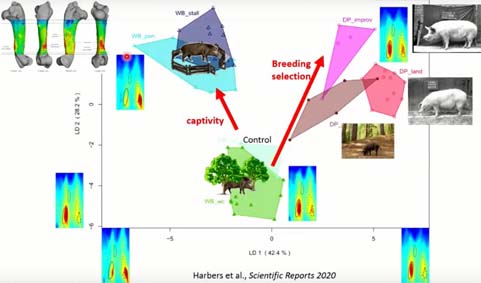
Si on compare les réponses entre captivité et sélection, une fois encore, on voit que la captivité entraîne une divergence morphologique dans la topographie de l’épaisseur corticale, mais toujours dans une direction différente de celle de la sélection. On voit une réduction des muscles adducteurs de l’épaule et une augmentation des muscles adducteurs du cou. On a des réponses claires et bien différenciées.
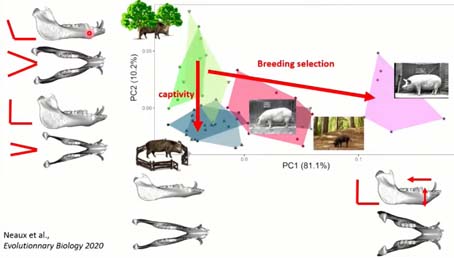
Pour le crâne, je ne vous présente que la mandibule, pour gagner du temps, mais les résultats sont cohérents aussi pour le crâne. On voit qu’ici aussi, avec la captivité, dans ce sens-là, la mandibule va changer de forme au cours de la vie des individus, avec des mandibules qui, par rapport au sanglier, ont un angle du ramus plus droit. La sélection entraîne elle aussi une réponse morphologique forte dans une direction, mais avec une certaine convergence avec également un angle de la mandibule qui devient plus droit.
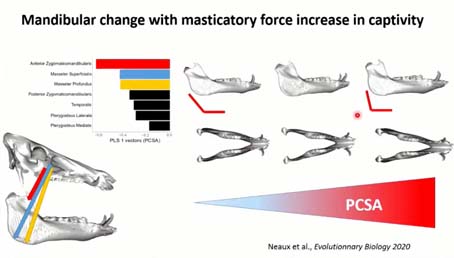
Cette convergence entre sélections et plasticité liée à la captivité, est liée à l’augmentation de la force musculaire, une fois encore, des individus en captivité, une augmentation de ces trois os qui relient le zygomatique avec la mandibule, qui en augmentant de force ont rendu l’angle de la mandibule plus droit.
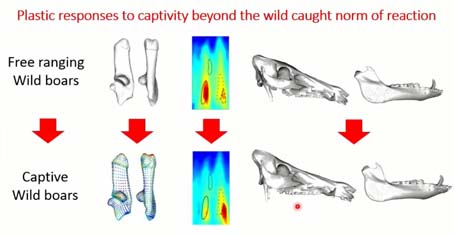
Pour terminer sur cette expérience, on a vu que quand on prend un animal sauvage et qu’on le fait grandir dans un environnement anthropisé où il ne peut plus exprimer l’ensemble de son répertoire comportemental, locomoteur, il y a un impact immédiat sur son squelette et sur sa forme crânienne. Cette différence se différencie bien de ce que l’on peut observer que dans les populations dans leur milieu naturel, mais aussi de la sélection des 200 dernières années.
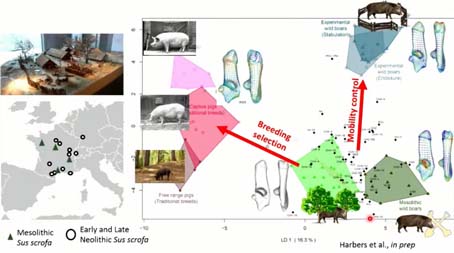
Si maintenant on regarde ce qui se passe dans le registre archéologique, on retrouve notre environnement des formes du calcanéum, cette fois-ci on a rajouté, ici, les sangliers du Mésolithique, les derniers sangliers chassés par les chasseurs du Paléolithique, on voit qu’ils ont quand même une forme assez différente de celle des sangliers actuels, et si l’on regarde l’endroit où cette différence s’exprime on peut considérer qu’il s’agissait d’individus plus âgés et peut-être beaucoup plus robustes.
Si on projette nos individus du Néolithique ancien et plus récent, on voit que leur distribution se situe dans le normes d’individus chassés mais quand même une distribution qui s’étire vers les individus dont le comportement a été plus ou moins contrôlé. Par contre, on n’a aucun individu tiré vers les phénotypes liés à la sélection des phénotypes.
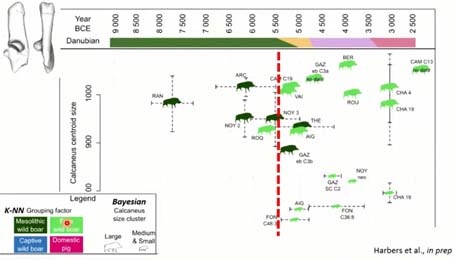
Maintenant, si on projette les résultats du calcanéum le long de la chronologie de nos sites étudiés, entre 8 000 et 2 500 avant Jésus-Christ, après les avoir analysés avec des approches bayésiennes, pour avoir des clusters de taille, et puis des analyses en machine learning, pour avoir des groupes phénotypiques, selon des groupes type-chassé et type plutôt sélectionné, on remarque que dans l’ensemble, tous les calcanéums archéologiques sont identifiés comme des sangliers dans leur milieu naturel, vraisemblablement. C’est vrai qu’on utilise des calcanéums, et pour qu’ils soient épiphysés, on a dû prendre des individus adultes. Si on avait eu de plus jeunes individus, peut-être qu’on aurait pu voir une … (phrase inachevée) Ce qui est intéressant ici, c’est qu’on a une augmentation de la taille des sangliers, et aussi, très intéressant, c’est qu’à partir de 5 000, on a l’apparition de petits phénotypes de sangliers. Est-ce qu’il s’agit de nouvelles populations allochtones ? Est-ce qu’il s’agit de sangliers, on va dire, commensaux, présélectionnés, donc un peu plus petit ? Ou est-ce qu’on a affaire à des populations hybrides ou férales ? Là, on a lancé des études moléculaires pour essayer d’apporter de nouvelles informations à ce sujet.
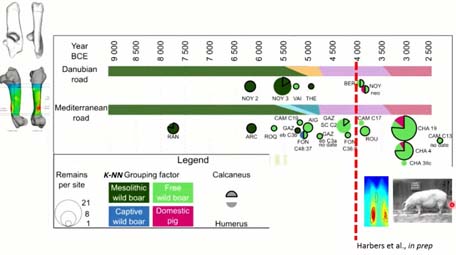
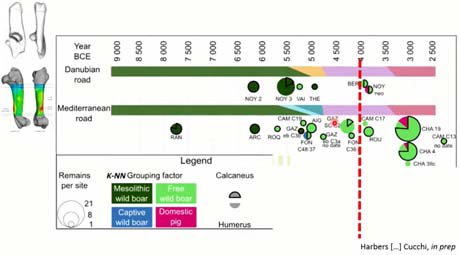
Si ensuite on cumul le calcanéum et l’humérus dans nos résultats, toujours sur cette chronologie, sur ces camemberts, la proportion avec le contour noir, c’est les résultats du calcanéums, et sans contour, il s’agit de l’humérus. Avec l’humérus on prend en compte une plus large gamme d’âges, donc on voit plus de choses, de façon anecdotique, on voit quelques individus captifs vers 5 000 et ce qui est intéressant, c’est qu’on voit des individus dans avec un phénotype sélectionné, types sélectionné, dès 4 000, que cela soit au Nord de la Loire ou dans la région de Jura, plutôt à l’Est de la France.
Évidemment, ce ne sont pas des phénotypes sélectionnés, tel qu’on le verra ces deux-cents dernières années, mais on a peut-être à faire aux premiers individus qui prennent plus de poids, qui sont plus lourds et par conséquent qui ont une allure un peu différente, et qui n’ont pas trop de différence de diaphyse.
Je vais terminer en disant, avant tout, un immense merci à tous mes collaborateurs, je pense particulièrement à l’équipe de la Réserve biologique de la Haute-Touche du Muséum d’histoire naturelle, qui ont fait un travail extraordinaire pour mettre en place l’expérimentation sur les sangliers, parce que, je vous assure, ce n’était pas une mince affaire. Et puis, bien sûr, je pense aux jeunes chercheurs, qui ont analysé et publié ces données. J’ai pensée particulière pour mon collègue Laurent PYMERAIL et ma collègue Anne TRESSET, qui nous ont quittés récemment au cours de cette recherche.
Voilà, merci beaucoup pour votre attention.
Frédéric DENHEZ : Merci à vous. J’avais juste une question : Est-ce que ce que vous avez observé est réversible ? Est-ce que les sangliers, une fois morphologiquement modifiés, si vous leu ficher la paix ils redeviennent, entre guillemets, « normaux » ?
Thomas CUCCHI : Oui, je pense que pour ce qui est des résultats que l’on a obtenus, si on laisse continuer à vivre encore deux ans mais cette fois dans un milieu totalement naturel, les marqueurs que l’on a mis en évidence auront disparus effectivement. Il ne s’agit là que de plasticité phénotypiques, il n’y a pas de mutations génétiques derrière. Il s’agit d’une réponse d’un génotype face à son environnement. Mais, ça, c’est ce que l’on va essayer de tester, il peut y avoir une empreinte épigénétique. Un empreinte environnementale sur le génome, qui pourra être transmise à la prochaine génération. Mais ça, c’est tout un pan, qu’avec mes collègues, on va essayer d’explorer.
Frédéric DENHEZ : Une question qui vient d’arriver sur internet : « Si vous aviez modifié la méthode d’« élevage », est-ce que cela aurait entraîné d’autres types de manifestations morphologiques ? » Autrement dit, est-ce qu’à un type d’élevage on peut avoir un type d’animal domestique ?
Thomas CUCCHI : Oui, oui, absolument, ça, c’est certain. C’est un petit peu ce qu’on observe. Maintenant, il y a des races un peu plus standardisées, ce que l’on appelait avant, pour le cochon, les large-white … interruption
Frédéric DENHEZ : Cela vous a tout figé. Vous avez parlé du cochon et cela vous fige … Thomas CUCCHI, on vous a perdu …
Monsieur ORLANDO, pouvez-vous répondre à la place de Thomas CUCCHI ? Si non, je fais une petite pause jusqu’à midi et quart. …
Je n’ai plus personne… Si, Ludovic ORLANDO ! Prenez la main, pour venir à l’aide de votre collègue.
Ludovic ORLANDO : J’étais un peu distrait, dites-moi.
Frédéric DENHEZ : La question est : Est-ce que, si on modifie les conditions de l’élevage, on aboutit à des modifications différentes du phénotype ?
Ludovic ORLANDO : Ça, c’est une très, très vaste question. En réalité, ce dépend du cadre temporel dans lequel on se place. Si l’on veut que la génétique soit un relais de cela, ça va dépendre immédiatement de l’intensité de la sélection que l’on fait. Pour caricaturer, imaginons qu’une maladie arrive dans un élevage, tue 99,999999% de l’élevage et qu’il n’y a qu’une survivante pour l’année d’après, à ce moment-là il y a une sélection très, très forte puisque ce n’est qu’elle qui conditionne la survie de ce qui se passe par la suite. Sinon, dans des phénomènes où c’est moins drastique, il va falloir un grand nombre de générations, en tout cas un certain nombre de générations pour que cela opère. Donc, la réponse à cela va dépendre de l’intensité de la sélection qui va être apportée, essentiellement de ces paramètres-là en réalité.
Frédéric DENHEZ : Merci, je fais une petite pause de cinq minutes, pour retrouver la table-ronde à midi et quart, pensez à …
Ah, Thomas CUCCHI, on vous a retrouvé, mais je suis désolée, je vous retrouve, et on vous retrouve tous ensemble, pour la table-ronde. Je fais une petite pause après vos deux exposés, brillants, roboratifs, ça nécessite cinq minutes de diffusion dans le cerveau avant d’aborder la table-ronde, à tout de suite.
Merci.