

Olivier BROSSEAU : Bonjour à tous.
Je vais être bref pour démarrer la matinée. Je vais me contenter de vous présenter Guillaume LECOINTRE, que certains d’entre vous connaissent. Il est professeur au Muséum national d’histoire naturelle, directeur du département Systématique et évolution. Dans le programme que vous avez reçu, son intervention avait pour titre : Comment définit-on l’évolution ? Il l’a renommée « Évolution, transformation ».
Évolution, transformation
Avec Guillaume LECOINTRE, Professeur du MNHN, Directeur du département Systématique & Évolution, Chef d’équipe à l’Institut de Systématique, Évolution, Biodiversité
Comme convenu avec les organisateurs des rencontres Culture en partage, mes collègues, qui interviendront tout au long de la journée, et moi-même avons bien pris en compte le fait que ces rendez-vous ne sont pas ceux de spécialistes en la matière, nous ne rendons pas compte ici de nos travaux d’universitaires. Je vais donc moi aussi centrer mon intervention sur ce que nous avons identifié comme obstacles pour les publics dans la réception de l’idée d’évolution.
Je vais vous parler de mon expérience de chercheur, engagé dans la diffusion et le partage des connaissances et savoirs, qui a observé des représentations et des blocages chez divers publics, sans doute les mêmes que vous retrouverez chez une partie des visiteurs de votre future exposition sur Darwin.
Thierry HOQUET l’avait dit lors de la séance précédente, l’évolution est un mot chargé de plusieurs sens. Quand vous parlez d’évolution au public, plusieurs images peuvent surgir dans l’esprit des uns ou des autres :
- D’abord, on peut entendre par évolution un processus par lequel les espèces se transforment. Je ne vais pas détailler le processus. Nous avons appris à l’école que l’évolution nous enseigne comment les espèces changent, évoluent au cours du temps. Nous verrons tout à l’heure que cette représentation mérite d’être pondérée. L’évolution, telle que nous la concevons aujourd’hui, explique autant la stabilité des régularités que nous voyons que les changements. Nous pouvons garder cette notion à l’esprit. J’y reviendrai dans un instant.
- L’évolution peut être comprise comme une théorie générale. La théorie générale de l’évolution est la théorie de la biologie, de l’anthropologie et de la paléontologie.
- L’évolution peut être conçue comme un déroulement historique des formes de vie au fil du temps à la surface de la planète. Cela ressemble à un scénario, qui raconterait l’Histoire du vivant. Cette représentation est extrêmement courante, peut-être l’image la plus populaire du mot évolution. Elle est conçue comme un déroulement de formes de vie au cours du temps.
- Sur le plan iconographique, l’évolution peut prendre l’image d’un arbre, qui se déploie. Cet arbre peut être représenté comme ici avec la racine en haut. Une figure montre des branches, qui représentent, dans la forme moderne, des degrés relatifs de parenté entre les espèces actuelles et fossiles.
- Pour finir, dans l’esprit de nos publics, l’évolution est aussi une marche vers le progrès, parce que nous avons un vieil héritage dans cette matière. Dans le vocabulaire courant, prendre quelqu’un à parti pour lui dire d’évoluer, cela signifie qu’il faut qu’il « se bouge ». Quand on dit de quelqu’un qu’il a évolué, en général, c’est un compliment. En revanche, dire que quelqu’un n’a pas évolué n’est pas flatteur. L’évolution est donc entendue comme un changement positif. Une mythologie du progrès a été associée à l’évolution dans la 2e moitié du XIXe siècle et au XXe siècle. Nous retrouvons cette marche vers un progrès chez Pierre-Paul Grassé, zoologiste français, auteur du traité de zoologie en 30 volumes, ou encore chez Jean Piveteau, ou Teilhard de Chardin. Le milieu du XXe siècle est donc également assez chargé de cette notion de progrès. Aujourd’hui, le point de vue est quelque peu différent.
À l’origine -Thierry HOQUET l’avait dit, je le rappelle - l’évolution dans les sciences n’était pas utilisée pour désigner ce que nous désignons aujourd’hui. L’évolution était l’équivalent de ce que nous appellerions aujourd’hui le développement embryonnaire : le déploiement en 3D de quelque chose déjà contenu en germe. Cet aspect représente un point de torsion important. Quand on s’intéresse à l’histoire des sciences, l’évolution, dans le passé (fin XVIIIe-début XIXe siècles), ne signifie pas du tout ce qu’elle signifie aujourd’hui. C’est même presque le contraire.
Si je vous dis que l’évolution sous-entend que ce qui est contenu en germe se déploie, cela signifie que ce qui se déploie est déjà prévu. Or, l’évolution, telle que nous la concevons aujourd’hui n’est pas prévisible sur le long terme : les milieux vont filtrer la variation spontanée et imprévisible manifestée par des individus et nous ne savons pas de quoi les milieux seront faits demain. L’évolution des populations, telle qu’elle est conçue aujourd’hui, ne déploie pas quelque chose contenu en germe.
Je ne suis peut-être pas exhaustif concernant les sens du mot évolution, mais au moins tous les sens que je viens de citer sont courants. Les scientifiques se concentrent davantage sur les 2 premiers. Dans les laboratoires, on appelle évolution tantôt le processus par lequel on comprend la transformation et le nom qu’on donne à la théorie générale de la biologie, de l’anthropologie et de la paléontologie.
Un problème d’enseignement et de communication avec le public sur l’évolution réside dans le fait que, si je vous dis que l’évolution est une théorie, les publics n’ont qu’une vision généralement péjorative de cette dernière. La théorie n’est pas conçue comme un élément solide en sciences. Le créationnisme, dont nous parlerons le 25 mars, utilise à plein le caractère quasi péjoratif du terme théorie. Malheureusement – je ne vous apprends rien –, on ne fait pas de science sans théorie. D’ailleurs, Whitman disait qu’« une théorie sans faits n’est que fantaisie, mais des faits sans théorie ne sont que chaos ». En science, les faits et les théories fonctionnent ensemble.
Les théories assurent la cohérence des faits entre eux et on change de théorie en choisissant la théorie la plus économique en hypothèses sur les faits. Moins il est nécessaire d’ajouter d’hypothèses surnuméraires non documentées, plus la théorie est cohérente. Plus il faut insérer des pièces de puzzles de force là où il n’y a pas de pièces, plus la théorie est faible. On préfère donc la théorie 1, où les pièces du puzzle s’agencent le mieux, à la théorie 2, où les pièces de puzzle n’entrent pas, et donc à cause de cela boursoufflée d’hypothèses non vérifiées. Le rôle d’une théorie consiste à agencer les faits, les lois, et des hypothèses. Plus une théorie est cohérente et plus elle rencontre de succès dans la prédiction des observations à venir. Le rôle d’une théorie est aussi de proposer des prédictions sur ce qui reste à observer et ne l’a pas encore été. Vous voyez la valeur considérable que peut revêtir la théorie en science.
Or, pour nos publics, les théories sont en général dévalorisées. Je ne sais pas pourquoi, mais c’est certainement lié à la façon, dont nous enseignons les sciences. Nous les enseignons comme des collections de faits, où seuls ces derniers seraient nobles. Nous avons un héritage de Cuvier. Nous avons un héritage de Claude Bernard. Chez Cuvier, nous remarquons un silence sur le rôle des théories en science. Les théories n’auraient été jugées, chez Cuvier, qu’en tant que spéculations.
Nos publics rencontrent également un problème cognitif sur l’évolution. L’évolution permet d’expliquer l’optimalité sans concepteur : une optimalité apparente, sans volonté. C’est-à-dire que nous sommes amenés à nous émerveiller de ce que j’appelle une optimalité, à savoir une apparente harmonie entre les formes et les fonctions dans la nature. Le terme harmonie relève toutefois du registre théologique, comme on en parlait au XIXe siècle.
Vous le savez, les organes nous semblent bien faits : « La courbure du cristallin, c’est juste ce qu’il faut pour que les rayons lumineux convergent sur la rétine : ah, que l’œil est bien fait ! » Nous avons une impression d’optimalité. Je dis bien une impression, dans la salle il y a des personnes qui portent des lunettes, cela veut dire que la courbure du cristallin n’a pas toujours la courbure qu’il faut pour que les rayons lumineux tombent sur la rétine. Nous sélectionnons les régularités, qui nous plaisent, pour penser que cette optimalité est normale. L’évolution vous apprend à obtenir cette optimalité sans concepteur. Nous passons notre temps à projeter nos propres réflexes dans la nature. Vous avez besoin de planter un clou dans un morceau de bois. Vous voulez fabriquer un marteau. Vous savez ce que devraient être les propriétés physiques de la tête et du manche du marteau, si vous voulez pouvoir vous en servir, et surtout comment associer le manche à la tête. Vous concevez. Vous anticipez les propriétés et les composantes de l’outil que vous allez créer.
Notre premier réflexe – vous le rencontrerez chez le public – est une transposition directe des réflexes cognitifs individuels dans la nature. Pourquoi l’évolution nous prend-elle à rebrousse-poil ? Elle nous raconte que nous avons des optimalités apparentes, mais sans conception. Cela est contre-intuitif, cela constitue un important obstacle à surmonter.
Évidemment, il existe la métaphore de l’essai-erreur. Pourquoi est-il difficile pour nous de considérer qu’une optimalité puisse être obtenue sans concepteur ? Nous oublions que les régularités du monde, qui nous plaisent, sont le fruit d’un jeu d’essai-erreur gigantesque. Pierre-Louis Moreau de Maupertuis, physicien du XVIIIe siècle avait été un des premiers à avoir cette intuition : est-ce que l’harmonie apparente dans la nature ne serait pas le fruit d’une vaste loterie ? Seules les quelques combinaisons propices à la vie auraient survécu et le reste serait mort et ne serait plus là pour qu’on en parle. Cette intuition du jeu d’essai-erreur est présente avant Darwin. De plus, elle n’est pas toujours remobilisée par les scientifiques.
Sur ce point, je vais laisser parler mes successeurs ce matin. Pour mieux comprendre le premier réflexe dans la tête des publics, si quelque chose apparaît dans la Nature, quand bien même accepterions-nous la transformation, que les espèces changent, chez nos publics elles changent parce qu’une volonté interne, une force vitale les pousse à changer. Une impulsion interne amène l’être vivant à manifester un changement, qui lui est favorable. C’est la figure du haut, raisonnement apporté en zoologie par Lamarck. Les êtres vivants, pour subvenir à leurs besoins, font des efforts. Ces efforts entraînent certains organes, c’est-à-dire les renforcent. Le défaut d’efforts sur d’autres organes diminue ces autres organes. Il y a donc des renforcements et des diminutions dans la vie de chaque individu, en vertu des besoins de l’organisme, qui sont transmis à la génération suivante.
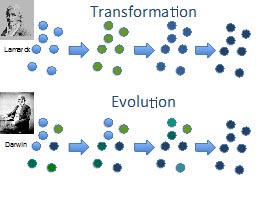
Les petites boules, que vous voyez, représentent les individus d’une population. Les corps physiques sont en quelque sorte moulés par l’environnement, imposant des contraintes, lesquelles imposent chez les organismes des efforts à faire pour subvenir à leurs besoins. C’est une théorie instructiviste du changement. Retenez bien dans ce schéma que ce qui se passe à l’échelle de l’individu reflète ce qui se passe à l’échelle de la population.
Deux propriétés essentielles différencient Lamarck de Darwin :
– La conception de la transformation est individuelle chez Lamarck (populationnelle chez Darwin). Si vous étudiez ce qui se passe chez l’individu, ce sera le reflet de ce qui se passe dans la population.
– Lamarck s’intéresse à la source de la variation, ce que ne fait pas Darwin. Les 2 auteurs ne regardent pas tout à fait la même chose au départ. Pour Lamarck, la variation apparaît aux besoins du porteur. Le changement (du bleu clair au foncé) apparaît aux besoins du porteur, puisque c’est en faisant des efforts que l’individu va renforcer ou diminuer ses organes.
J’ai cité deux différences majeures avec Darwin. Dans la figure du bas, vous remarquerez que la variation est déjà là. Darwin ne s’intéresse pas directement à la source de la variation. Il lui suffit de constater qu’elle est là. Il démarre avec la variation comme fait premier. Il ignore pourquoi il y a tant de variations. Plus précisément, il a quelques idées sur les causes globales des variations en général, mais il ignore tout de l’apparition d’une variation particulière. Ce qui va compter pour lui, c’est ce qui va se passer après. Ainsi, le changement ne se manifeste pas forcément en vertu d’efforts, mais dans toutes les directions. Cela constitue une différence majeure. Chez Darwin, la variation n’apparaît pas forcément en prévision des besoins de celui qui la porte. Elle peut donc être néfaste, avantageuse ou neutre pour le porteur. Seules les variations avantageuses pour le porteur en termes de nombre de petits vont avoir tendance à s’installer dans la population. Un changement obtenu par voie de sélection naturelle se manifeste ainsi.
S’agissant de la sélection naturelle, on insiste souvent sur son rôle pour expliquer le changement, mais finalement, quand le milieu est relativement stable, ce processus explique pourquoi les mêmes variantes se maintiennent dans une forme régulière. Le processus explique aussi la régularité. Or, on oublie souvent de souligner ce point dans l’enseignement, et il y a des conséquences majeures à cela. Le principe de sélection naturelle, que vous voyez en bas, explique autant le changement quand le milieu change qu’il peut expliquer le maintien lorsque le milieu ne change pas. Le frontispice de l’Origine des espèces parle de « préservation » des espèces avantagées, non de changement. Le mot évolution n’est même pas dans le titre du livre, pour des raisons qui vous ont été expliquées la dernière fois.
Nous reviendrons assez longuement sur la représentation du public sur ces deux processus. Retenez simplement que ce qui fait la différence entre le haut (Lamarck) et le bas (Darwin), n’est pas tant lié à l’héritabilité des caractères acquis au cours de la vie individuelle. À son époque, Darwin admet tout à fait l’héritabilité des caractères acquis au cours de la vie individuelle. Ce critère ne fonde pas la différence entre les deux auteurs. Ceux-ci diffèrent beaucoup plus par les deux paramètres, que je viens de citer : premièrement la variation apparaît selon les besoins de son porteur chez Lamarck, et indépendamment des besoins du futur porteur chez Darwin ; deuxièmement la transformation est individuelle chez Lamarck et populationnelle chez Darwin.
Je souhaite vous alerter sur un piège, qui réside dans la réduction des effets de la sélection naturelle. Quand nous parlons de sélection naturelle dans les journaux, notamment ceux de l’économie, le premier élément souligné est la compétition entre les individus pour la captation des ressources. Nous avons gardé cet élément d’une formule de communication assez concise, le fameux struggle for life. En français, struggle ne se traduit pas par combat, mais plutôt par lutte. Quand vous parlez de lutte, cette métaphore est extrêmement large. Vous pouvez lutter contre le froid. Toutes les actions que les organismes sont amenés à exécuter dans leur vie quotidienne sont des efforts, qui peuvent être mis sous le chapitre de la lutte. Coopérer avec son voisin relève de la lutte, mais d’une lutte collective et non individuelle. Vous devez comprendre la métaphore de la lutte sous un aspect extrêmement large. La « lutte pour l’existence » inclut donc la coopération.
Or, dans les journaux, nos publics sont confrontés à une réduction systématique de la sélection naturelle soit (1) à la compétition entre les individus pour la captation des ressources ou (2) au camouflage (exemple des fameux papillons de Kettlewell : la phalène du bouleau). Quand on essaie de donner un exemple de sélection naturelle, on montre des organismes pris dans un combat singulier, ou qui en ressemblant à leur milieu, échappent à leurs prédateurs. La vraie question pour comprendre la richesse des conséquences de la sélection naturelle est de savoir quelles sont les dispositions ou les capacités permettant aux porteurs de produire plus de petits lors de la génération suivante. Si vous posez la question de cette manière, vous allez vous apercevoir que cela n’englobe pas seulement la compétition et le camouflage.
Le camouflage est un concept relativement facile à comprendre. Les traits, acquis par hasard, qui permettent aux porteurs de mieux ressembler à leur milieu, vont avoir tendance à se stabiliser dans la population s’ils sont héritables. En effet, en ressemblant davantage au milieu, on échappe aux prédateurs, et on a plus de chances de faire de petits. Il y a de plus un effet cumulatif (en nombre de petits laissés dans les générations suivantes) des variants positifs pour le porteur, tandis qu’il n’y a pas d’effet cumulatif pour les variations neutres ou néfastes. D’où le renforcement mutuel des traits avantageux, qui nous laisse l’impression que la population est adaptée et que des individus sont « bien faits ».
Deuxième conséquence sélective : la capacité des individus à capter davantage de ressources. Ceux des variants, qui par hasard permettent de capter de la ressource, s’ils sont héritables, vont avoir tendance à se répandre dans les populations. C’est une des raisons pour lesquelles il y a une aptitude à capter de la ressource, mais également une lutte interindividuelle pour préserver cette ressource. On peut citer les renards ou encore les aigles, qui se réservent des territoires de 5 km2 de chasse, où ils sont les seuls à chasser, en évacuant tout congénère, sauf les femelles au moment de la reproduction.
Justement, parlons de la reproduction, non pas chez Darwin, mais dans la biologie contemporaine, une des conséquences sélectives est aussi la capacité des individus à capter l’attention du sexe opposé. Vous comprenez pourquoi. En effet, si ce qui compte en termes de sélection est le nombre de petits à la génération suivante, portant vos propres caractéristiques. Or, l’attraction sexuelle est la clé de l’accès à la production de la génération suivante. Il est donc très important de séduire le sexe opposé. Cet aspect de la sélection naturelle s’appelle la sélection sexuelle.
Cependant, Darwin ne plaçait pas la sélection sexuelle sous le chapitre de la sélection naturelle. Toutefois, la science d’aujourd’hui considère que la sélection sexuelle est une des modalités d’une sélection naturelle plus largement comprise.
Typiquement, ce paon mâle présente une traîne magnificente. Il a été expérimentalement montré que celle-ci a un rapport avec le succès du porteur, lorsqu’elle est déployée, à attirer le consentement à l’accouplement par la femelle.
Je vous parle de ces conséquences sélectives, car j’aimerais vous sensibiliser à une idée, qui m’est chère. Vous comprenez tout de suite que ces différentes catégories de conséquences ne sont pas toujours cohérentes entre-elles et cela est très important à comprendre. La sélection naturelle ne fabrique pas de perfection. Tout ce que l’on voit dans la nature, obtenu par sélection naturelle n’est pas forcément adapté. Tout est une question de degrés, il y a des compromis. Ainsi, ce paon est peut-être efficace pour séduire la femelle, mais il le sera moins au moment de fuir un prédateur.
En biologie des populations, aujourd’hui, on calcule les bénéfices et les coûts, la métaphore n’est pas très jolie, mais il y a ce que l’on appelle des fardeaux. C’est-à-dire que des organes se mettent en place, mais sont coûteux, en termes de descendance, car il y a des tensions sélectives contradictoires sur un même organisme. Nous avons du mal à expliquer cette notion au public, car l’idée de perfection, qui n’est pas une notion scientifique, reste sous-jacente. La perfection biologique n’existe pas.
Une autre conséquence sélective se trouve dans les associations d’espèces. C’est ce qu’on appelle la symbiose, c’est-à-dire le vivre ensemble. La symbiose est absolument générale dans le vivant. Il n’existe pas un être vivant, qui ne soit pas symbiotique. Rappelez simplement au public qu’il ne pourrait pas vivre sans sa flore intestinale par exemple. Celle-ci comporte mille fois plus de cellules qu’il n’a de cellules propres. Nous sommes des fermenteurs à bactéries. La symbiose est absolument généralisée. Retenez sur ce plan, qu’il y a un continuum entre des situations où ces associations sont à bénéfices déséquilibrés, et les situations où les bénéfices sont équilibrés.
Les premières sont appelées parasitisme, les secondes sont appelées mutualisme. L’image forte illustrant cette situation de mutualisme est celle de ce zèbre, qui laisse les pique-bœufs picorer sur son pelage. Le zèbre se débarrasse ainsi des parasites sur son pelage, tandis que le pique-bœuf se nourrit. Le parasitisme, quant à lui, est généralisé dans tout le vivant : toute espèce constitue une ressource permanente pour au moins une autre. Nous l’oublions un peu vite, parce que nous vivons dans des conditions d’hygiène exceptionnelles, mais souvenons-nous que nous avons eu des poux ou des puces au moins une fois dans notre vie.
Nous avons parlé de mutualisme, c’est-à-dire d’entraide interspécifique, comme étant l’une des modalités de la symbiose, mais vous avez aussi de l’entraide intraspécifique à l’origine des comportements sociaux dans bien des lignées de vivants (insectes, mammifères). Cette idée renvoie à la mutualisation des bénéfices. En effet, on s’en sort mieux en termes de nombre de petits, lorsqu’on coopère face aux aléas de l’environnement, à l’échelle collective qu’on ne s’en sortirait chaque individu, si chacun restait de son côté. La métaphore qui illustrerait le mieux cette situation serait « l’union fait la force ».
Les lycaons présentent une réelle mutualisation de l’effort de chasse. Aucun des lycaons ne pourrait seul tuer un gnou ou un zèbre. Ils y parviennent, parce qu’ils chassent collectivement à l’endurance. L’animal est dépecé sur place à plusieurs dizaines de kilomètres de la tanière. La viande est ramenée à la tanière. Chaque individu régurgite de la viande pour tous les petits restés à la tanière, indistinctement. Les individus régurgitent également pour ceux des adultes restés à la tanière pour garder la crèche. Ce ne sont jamais les mêmes individus, qui gardent la crèche d’une chasse à l’autre. Bref, l’effort de chasse est ainsi mutualisé. Nos sociétés sont des compromis entre des comportements égoïstes (compétition pour la captation des ressources) et d’efforts solidaires.
L’idée que la sélection naturelle mène à des compromis et non à des perfections doit être retenue. Elle permet de lutter contre la représentation courante, qui consiste à dire que la sélection naturelle organise tout pour le mieux. Je conclurai en soulignant que l’idée d’évolution renvoie essentiellement à la sélection naturelle. Celle-ci explique autant la régularité de ce que nous voyons que son changement.
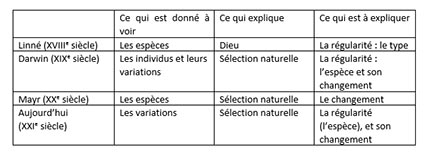
Dans le monde de Linné, botaniste Suédois, Dieu explique la régularité de ce qui est donné à voir, à savoir les espèces. Elles sont l’entité première qui s’offre à l’œil de l’humain. Elles sont repérées par un type. Pourquoi parlons-nous encore de Darwin de nos jours ? Il a été le premier à inverser la vapeur, car il étudie en premier lieu les individus et leurs variations, et don des espèces. Les espèces, il en cherche l’origine ! Je pourrais être plus long sur cette question. La sélection naturelle explique d’abord la régularité des individus qui font des petits ensembles. Quand ces individus sont réguliers, nous les appelons espèces. Une espèce n’est en somme qu’une convention de langage posée sur une hypothèse ou un fait d’isolement reproducteur.
Quand on parle de sélection naturelle, on sélectionne quelque chose, on maintient certaines dispositions. Dans l’Origine des espèces, de Charles Darwin, ce qui est amusant à considérer, c’est l’origine de l’espèce en tant que régularité physique et non en tant qu’entité créée dans la nature de toutes pièces. On remarque que le sous-titre de l’Origine des espèces parle de « préservation » et non d’évolution. Ainsi, la sélection naturelle explique d’abord pourquoi ce que vous observez globalement est régulier malgré l’irrégularité du changement permanent des individus. Les individus changent et varient et la sélection naturelle vous explique comment, dans un milieu stable, il y a quelque chose de régulier, alors qu’à petite échelle, tout est irrégulier.
Ce dont je vous parle ici est le contraire de ce qu’on m’a enseigné. Suite à l’influence d’Ernst Mayr, à l’école, on m’avait appris que la sélection naturelle expliquait le changement des espèces. En effet, pour Ernst Mayr, l’espèce était première. Dans Principles of Systematic Zoology en 1969, Ernst Mayr pratique un réalisme de l’espèce. C’est-à-dire que l’espèce est première à voir. Dès lors, le rôle de la sélection naturelle est d’expliquer le changement de quelque chose de régulier de fait.
Or Darwin nous explique comment, grâce à la sélection naturelle, vous obtenez une régularité, qui n’est pas évidente sur une base de changement permanent. On nous a enseigné à l’école une évolution renversée comme une chaussette par rapport à l’évolution de Darwin. Attendez-vous donc à ce que le public pense d’abord que la sélection naturelle ait pour rôle d’expliquer le changement. Le public, qui lit de l’Origine des espèces de Darwin, pensera que ce livre ne traite pas de l’origine des espèces. Au XXIème siècle, comme l’espèce n’est pas donnée à voir, mais qu’elle représente une convention de langage posée sur une régularité isolée du point de vue de la reproduction, la sélection naturelle explique comment vous rencontrerez quelque chose de régulier à grande échelle à partir des individus et leurs variations qui se manifestent à petite échelle.
Comment les individus qui constituent les générations restent-ils réguliers malgré le changement ? Voilà le rôle de la sélection naturelle. Ensuite, si le milieu change, évidemment la sélection naturelle va vous expliquer comment et pourquoi les populations et les espèces changent. Attention, je n’ai pas dit que la sélection naturelle n’expliquait pas le changement. Elle l’explique bien sûr, mais, en première instance, elle est faite pour expliquer pourquoi il subsiste une régularité malgré le changement physique des individus. Quand le milieu change, elle explique aussi comment la population change.
J’en ai fini. Je suis prêt à répondre à vos questions. Merci.
Applaudissements
Olivier BROSSEAU : Merci Guillaume. Je vous propose de prendre une ou deux questions avant d’écouter Laurent LOISON, pour l’intervention autour de Lamarck et Darwin.
Vous nous avez parlé de prédiction, au début de votre intervention, peux-vous nous donner quelques précisions et exemples de prédiction ?
Guillaume LECOINTRE : Oui, il y en a un que j’aime particulièrement. Toutefois, il convient de distinguer les prédictions de long terme et de court terme. Les prédictions de long terme frappent davantage le public.
Concernant les degrés relatifs d’apparentement, construits à partir d’arbres, nous avions récemment découvert, grâce à la comparaison du matériel génétique, que les cétacés (les baleines et les dauphins) tiraient leur origine au beau milieu des vaches et des hippopotames (les artiodactyles). Il y a avait longtemps que les paléontologues réfléchissaient sur l’origine des baleines. Les cétacés sont nichés en plein milieu des animaux à deux sabots, les artiodactyles.
À quoi reconnaît-on un artiodactyle dans la nature ? Dans la cheville, il dispose d’un os, appelé l’astragale. Quand on joue aux osselets, on joue avec des astragales de mouton. Il y a une poulie dessus et dessous, c’est donc un astragale à double poulie. Il s’agit de la signature phylogénétique des artiodactyles. Or, du temps où les ancêtres des baleines avaient des pattes (il y a -50 ou -40 millions d’années), ces « baleines » ont dû avoir un astragale à double poulie, puisqu’elles émanent de la souche des artiodactyles. Cependant, nous ne disposions d’aucun modèle de baleine à pattes. Personne ne pouvait donc pas vérifier cette prédiction.
Pourtant, on a découvert un fossile dans l’Éocène au Pakistan au début des années 1990. Ce fossile était celui d’une « baleine » à pattes, présentant tous les caractères des cétacés et des membres postérieurs. Malheureusement, le fossile était coupé juste avant la cheville. Les paléontologues enragèrent de frustration, car nous ne saurions pas si ce fossile était doté d’un astragale à double poulie. Or, quelques années plus tard, on a retrouvé dans la même région un autre fossile doté d’un crâne de « baleine », dont les membres postérieurs étaient fossilisés. Effectivement, il avait bien un bel astragale à double poulie à la cheville.
Les phylogénies, c’est-à-dire ces arbres qui vous expliquent qui est apparenté à qui, permettent en partie de réaliser des prédictions. Celles-ci n’ont rien de formel. Elles se gèrent en termes de probabilités. Ce n’est pas une loi, mais il reste probable de considérer que les événements se soient déroulés de telle manière (raisonnement adductif). Parfois, il est même possible de le vérifier.
Régis MEYRAN : Ma question visait à rebondir sur une remarque, que je trouve assez difficile à comprendre, sur le fait que l’espèce est vue avant tout comme une convention de langage. Pour faire le lien avec des questions très concrètes des publics de la Cité des sciences notamment, j’aimerais aborder la question des races humaines. Celles-ci sont classiquement considérées comme une sorte de sous-espèce quelque peu floue, mais qui existe notamment chez Buffon et chez un certain nombre de naturalistes.
Il y a eu beaucoup de débats autour de la réalité ou non de cette notion de races humaines. Je voulais avoir votre avis, sachant qu’un des éléments avancés pour critiquer cette notion est de souligner qu’une des caractéristiques des êtres humains depuis leur apparition en Afrique est la dispersion géographique, qui aurait abouti au fait que la sous-espèce n’aurait pas eu le temps de se fixer précisément.
Si l’espèce est une convention de langage, qu’en est-il de la sous-espèce ?
Guillaume LECOINTRE : Mon métier est de faire des catégories, je suis systématicien de métier. Il existe mille façons de découper le monde. La manière que nous avons de le découper résulte d’une convention. Découper le monde naturel en entités généalogiques est une décision prise au chapitre 13 de L’Origine des espèces de Darwin. La classification légitime va être celle, qui tient compte des apparentements. Ainsi, la façon, dont nous découpons les espèces, les genres, les familles et les ordres, résulte d’abord de l’intention du classificateur en premier lieu.
Pour en venir à l’espèce, l’espèce renvoie à un découpage du monde marqué par un élément réel, à savoir une barrière à la reproduction. Des barrières à la reproduction existent vraiment dans la nature. En revanche, la boîte que je vais fabriquer à partir de là reste un élément intellectuel. La dernière fois, j’avais choisi la métaphore de Linné qui avait fait les pachydermes. Le critère justifiant la catégorie de pachydermes était lié à la peau épaisse. Dans la même boîte étaient donc rangés des éléphants, des hippopotames et des rhinocéros. Aujourd’hui, la boîte pachyderme a disparu de la zoologie. On ne parle plus de pachyderme. Pour autant, les hippopotames, les éléphants et les rhinocéros n’ont pas disparu. On ne confond donc pas la validité du contenant et l’existence matérielle du contenu. Les pachydermes avaient la peau épaisse, et c’est la raison pour laquelle le groupe avait été fait. Ils l’ont toujours. Le groupe n’existe pourtant plus, parce qu’il est incohérent du point de vue de l’apparentement. Et que cet apparentement est le critère de validité des groupes retenu aujourd’hui. Ainsi, il y a des caractères, qui existent, et des taxons, qui éventuellement n’existent plus. Analogiquement, chez l’homme on peut détecter des caractères raciaux, comme la couleur e la peau, mais cela ne signifie pas que les races sont valides. Souvenez-vous : il n’y a plus de pachydermes, mais les rhinocéros ont toujours la peau épaisse ! Venons-en aux espèces. Dans le monde matériel, il existe des barrières à la reproduction. À partir de là, j’instaure une barrière conventionnelle dans mon esprit, qui s’appelle l’espèce. Cette boîte vous ne l’aurez jamais dans sa totalité et vous ne pourrez la concevoir qu’avec la profondeur du temps. Nous reviendrons plus tard sur cela, si vous le souhaitez. C’est le sens de mon propos, quand je dis que l’espèce est conventionnelle. L’espèce reste un nom posé sur une boîte que nous avons fabriquée, et qui désigne une hypothèse de communauté de reproduction (que je n’observe jamais dans sa totalité). Je vous garantis que, quand on divise une espèce en deux ou quand on associe des populations, qu’on croyait séparées, dans une même espèce, il ne se produit rien dans le monde réel.
Il en va de même pour la sous-espèce. Elle relève d’une convention. Quel est le critère pertinent pour diviser une espèce en plusieurs sous-parties ? Par convention, nous instaurons des sous-espèces phylogéographiques. Dans ce cas, il est bon de donner le critère ayant servi à les distinguer.
Vous ne portez pas une essence de la sous-espèce en vous. Simplement, sous des critères phylogéographiques, on vous a donné un nom différent, qui vous fait appartenir à la race x ou y. Par ailleurs, il convient d’employer un autre mot que le mot race, très connoté, et surtout utile en zootechnie, où l’homme maîtrise les flux reproductifs. Vous pouvez parler de sous-espèces ou de groupes phylogénétiques. En tout cas, nous ne pouvons pas nier qu’il reste, dans la diversité humaine, des traces des mouvements de populations au cours des 120 000 ans d’histoire de l’espèce humaine. Reste à savoir si les caractères qui témoignent de cette histoire sont assez cohérents entre eux pour que nous puissions forger une « boîte » (un taxon de rang sous-espèce ») assez solide. Or, ces caractères ne le sont pas : par exemple, les groupes sanguins ou la taille ne se corrèlent pas à la couleur de la peau. C’est pour cela que l’on dit qu’il existe des caractères raciaux particuliers, mais que les races n’ont pas de cohérence.
Olivier BROSSEAU : Je propose de passer à l’intervention de Laurent LOISON, qui est historien de la biologie à l’université de Strasbourg et qui va nous parler de Darwin et de Lamarck.
Applaudissements